
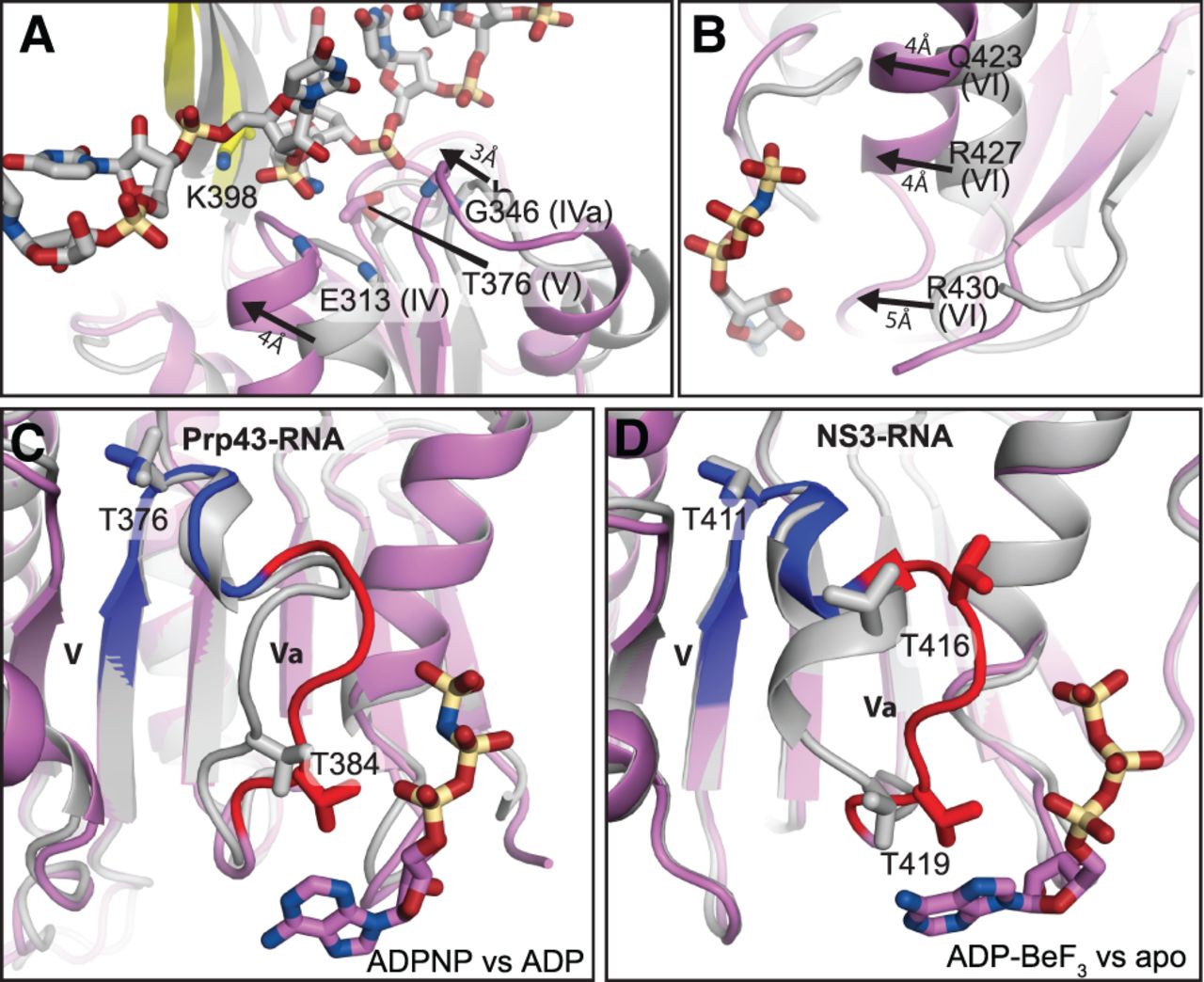
The rotation of RecA2 enables RNA and ADPNP binding, and alternative motif Va conformations implicate this element as an ATP sensor. (A,B) The 9° rotation of RecA2 enables binding of both RNA (A) and ATP (B), implicating a mechanism by which RNA binding stimulates ATPase activity. In panel A, the movement of motifs IV and IVa in the RNA- and ADPNP-bound structure, relative to the ADP-bound structure (PDB 3KX2), is indicated by arrows. In panel B, the movement of motif VI in the RNA- and ADPNP-bound structure, relative to the ADP-bound structure, is indicated by arrows. In panels A and B, the RecA1 domain of Prp43p in the RNA-free, ADP-bound (gray) and the RNA- and ADPNP-bound (colored) states were superimposed. (C,D) In Prp43, as in NS3, motif Va rearranges, and movement correlates with loss of the γ-phosphate. Panel C compares the ADPNP- and RNA-bound state versus ADP-bound state of Prp43p; panel D compares the ADP–BeF3- and RNA-bound state versus the apo, RNA-bound state of NS3 (PDB 3O8R versus 3O8C). In each case, the RecA2 domain of the ATP-bound state (colored) was superimposed with the RecA2 domain of the contrasting states (gray). Blue highlights the static nature of motif V while red highlights the dynamic nature of motif Va. The ATP analog is shown, colored by atom type. RNA is omitted for clarity.








