
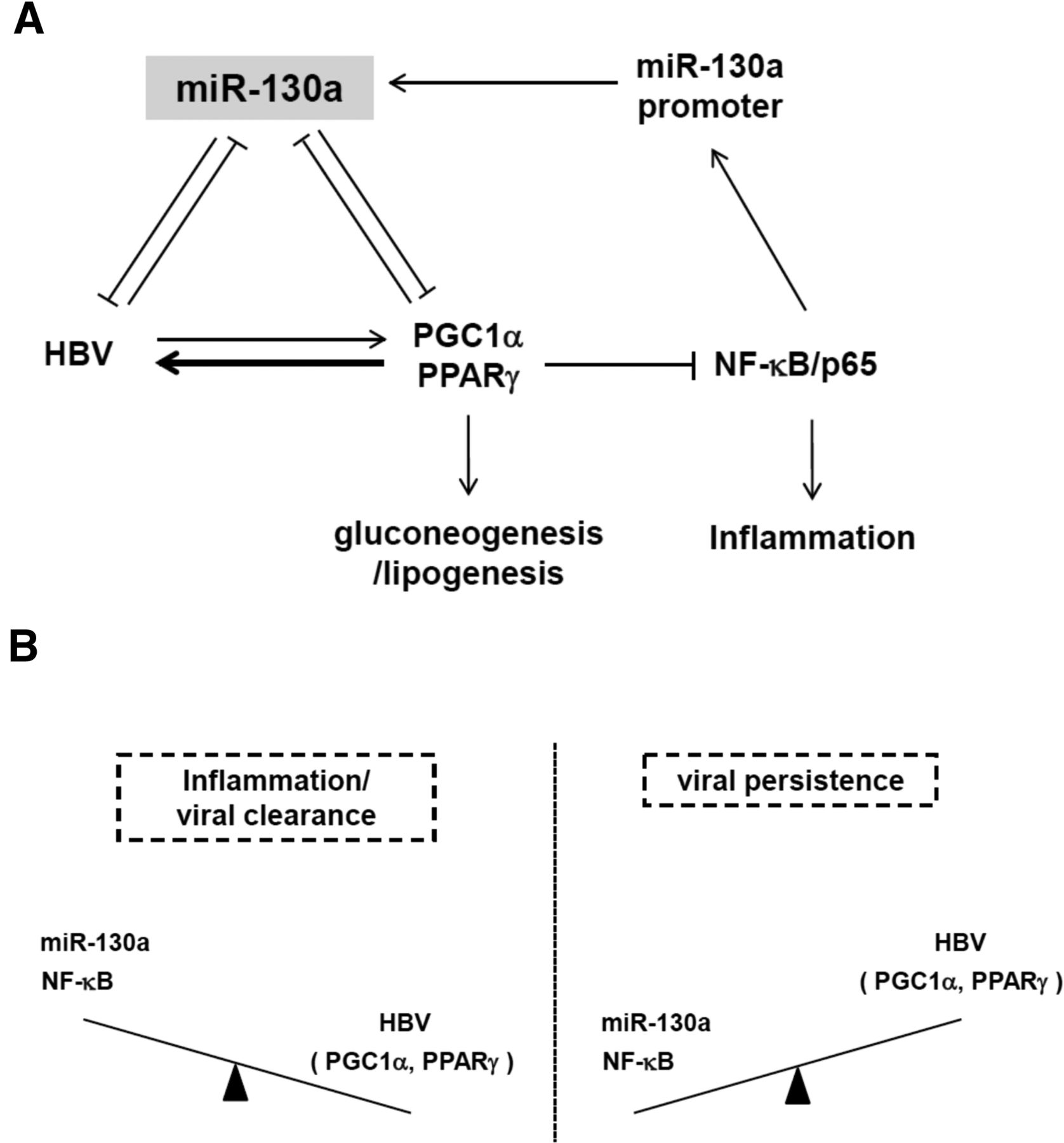
Diagrams of virus-host interactions between HBV and miR-130a, NF-κB, PGC1α, and PPARγ. (A) A cartoon summarizes the integration of two positive feed-forward loops from Figures 7I and 6C. In the presence of HBV, the level of miR-130a was reduced (Table 1; Fig. 1A), probably due to a reduced level of NF-κB/p65 in hepatocytes (Figs. 7G, 9C). Since miR-130a can target both PGC1α and PPARγ mRNAs simultaneously (Figs. 3C,D, 6C), the reduction of miR-130a can result in elevated levels of PGC1α and PPARγ, which can coactivate HBV transcription, leading to increased HBV DNA replication (Fig. 5B). It has been reported previously that PPARγ protein can serve as an E3 ligase for NF-κB/p65, resulting in NF-κB/p65 protein ubiquitination and degradation, leading to the reduction of NF-κB/p65 and miR-130a promoter activity (Fig. 7I). In this scheme (A), HBV can create a more friendly niche for itself by reducing the level of miR-130a via a positive feed-forward loop (Fig. 7I). Metabolically, PGC1α is known to be a positive transcriptional coactivator of hepatic gluconeogenesis (Yoon et al. 2001). PPARγ is a positive transcription factor for lipogenesis which in turn can reduce blood glucose level (Yu et al. 2003; Liu et al. 2012). By dual targeting at PGC1α and PPARγ, miR-130a could play a critical role in glucose homeostasis. (B) Liver inflammation could favor viral clearance, since when NF-κB and miR-130a are elevated, PGC1α, PPARγ, and HBV replication are reduced. In contrast, when liver is without inflammation, the levels of both NF-κB and miR-130a are low, and the levels of PGC1α and PPARγ are higher, resulting in more active viral replication.








