
Strict 3′ splice site sequence requirements for U2 snRNP recruitment after U2AF binding underlie a genetic defect leading to autoimmune disease
Abstract
We report that the 3′ splice site associated with the alternatively spliced exon 6 of the Fas receptor CD95 displays strict sequence requirements and that a mutation that disrupts this particular sequence arrangement leads to constitutive exon 6 skipping in a patient suffering from autoimmune lymphoproliferative syndrome (ALPS). Specifically, we find an absolute requirement for RCAG/G at the 3′ splice site (where R represents purine, and / indicates the intron/exon boundary) and the balance between exon inclusion and skipping is exquisitely sensitive to single nucleotide variations in the uridine content of the upstream polypyrimidine (Py)-tract. Biochemical experiments revealed that the ALPS patient mutation reduces U2 snRNP recruitment to the 3′ splice site region and that this effect cannot be explained by decreased interaction with the U2 snRNP Auxiliary Factor U2AF, whose 65- and 35-kDa subunits recognize the Py-tract and 3′ splice site AG, respectively. The effect of the mutation, which generates a tandem of two consecutive AG dinucleotides at the 3′ splice site, can be suppressed by increasing the distance between the AGs, mutating the natural 3′ splice site AG or increasing the uridine content of the Py-tract at a position distal from the 3′ splice site. The suppressive effects of these additional mutations correlate with increased recruitment of U2 snRNP but not with U2AF binding, again suggesting that the strict architecture of Fas intron 5 3′ splice site region is tuned to regulate alternative exon inclusion through modulation of U2 snRNP assembly after U2AF binding.
Keywords
INTRODUCTION
Removal of introns from pre-mRNA precursors (pre-mRNA splicing) is a necessary step for the expression of the majority of higher eukaryotic genes (Wahl et al. 2009), and alternative patterns of intron removal greatly expand the information content of complex genomes (Nilsen and Graveley 2010). Three sequence elements help to specify the 3′ end of introns in higher eukaryotes: the branch point, the 3′ splice site AG, and the intervening polypyrimidine (Py)-tract. The branch point, a virtually invariant adenosine that forms a 2′-5′ phosphodiester bond with the 5′ end of the intron after the first catalytic step of the splicing reaction, is flanked by the sequence UACUAAC in budding yeast (A denotes the branch point adenosine) and by the much more variable consensus YNYURAY in human genes (Y indicates pyrimidine; R, purine; and N, any nucleotide) (Zhang 1998). The branch point region is first recognized by the Branch Point Binding Protein (BBP/SF1) (Berglund et al. 1997; Liu et al. 2001) and subsequently through base pairing interactions with U2 snRNA, the RNA component of the U2 snRNP ribonucleoprotein (Nelson and Green 1989; Wu and Manley 1989; Zhuang and Weiner 1989). The invariant 3′ terminal AG dinucleotide and the preceding Py-tract are recognized, respectively, by the 35- and 65-kDa subunits of the U2 snRNP Auxiliary Factor (U2AF), which assists in the recruitment of U2 snRNP to the branch point region (Zamore et al. 1992; Merendino et al. 1999; Wu et al. 1999; Zorio and Blumenthal 1999).
A large variety of sequences fit into this general organization of the intron 3′ end, and cooperative interactions help to functionally define 3′ splice sites. For example, interaction between U2AF and BBP/SF1 (Selenko et al. 2003) mutually stabilizes the binding of these factors to their loosely defined adjacent cognate sites (Berglund et al. 1998). Another example is stabilizing interactions mediated by arginine-serine–rich (RS) domains, which support protein–protein interactions as well as interactions between proteins harboring these domains and double-stranded RNA regions (e.g., those resulting from base pairing between U2 snRNA and the branch site region) (Wu and Maniatis 1993; Shen and Green 2004 2006; Shen et al. 2004). Furthermore, the arrangement of protein and RNA contacts in the initial complex has been proposed to enforce a unique RNA structure in the 3′ splice site that may be critical to trigger subsequent events in the splicing process (Kent et al. 2003). In addition, proofreading activities exist that ensure proper assembly of splicing factors in bona fide splice site signals, e.g., allowing the assembly of U2AF on uridine-rich sequences only if followed by a 3′ splice site AG dinucleotide (Soares et al. 2006).
A third mechanism contributing to splice site identification is the sequential recognition of the same signal by different factors at different stages of spliceosome assembly or during the processes leading to catalysis. For example, the 3′ splice site AG dinucleotide is recognized initially by U2AF35, an interaction that becomes essential for spliceosome assembly on AG-dependent introns, which are characterized by relatively uridine-poor Py-tracts that can only weakly recruit U2AF65 in the absence of the U2AF35/AG interaction (Wu et al. 1999). The splicing factor hSlu7 restructures the spliceosome after the first catalytic step of the splicing reaction and positions the 3′ splice site AG for engaging in the second catalytic step over neighboring AG dinucleotides, thus helping to generate properly spliced mRNA products (Chua and Reed 1999a,b). Neighboring AGs can, however, strongly influence splice site choice, facilitating early interactions with the spliceosome but allowing other sites to be used during catalysis (Chua and Reed 2001; Lallena et al. 2002; Lev-Maor et al. 2003). 3′ splice sites are further recognized by proofreading mechanisms implemented in budding yeast by the DExD/H-box ATPase Prp22p that prevent exon ligation in pre-mRNAs containing mutations in splicing signals, including the 3′ splice site (Mayas et al. 2006).
The Fas/CD95 gene encodes a pro-apoptotic membrane receptor (Strasser et al. 2009) that has been recently shown to be also required for efficient growth of cancer cells (Chen et al. 2010). An alternatively spliced variant that skips exon 6 lacks the trans-membrane domain and encodes a soluble form of the receptor that can be anti-apoptotic (Cheng et al. 1994). A switch in alternative splicing of exon 6 occurs during T-cell activation such that the anti-apoptotic form of the receptor is produced during antigen-triggered clonal amplification, while the pro-apoptotic form contributes to eliminate the expanded cell population upon completion of the immune response (Liu et al. 1995; Russell 1995). Failure to accomplish this alternative splicing switch is linked with persistent high levels of lymphocytes in the lymph nodes, liver, and spleen, which lead to autoimmune lymphoproliferative syndrome (ALPS) (Roesler et al. 2005). Several splicing factors have been shown to modulate the levels of Fas exon 6 inclusion or skipping by various molecular mechanisms, including targeting early events in splice site recognition/exon definition and modulating splice site pairing after U2 snRNP binding (Förch et al. 2000; Izquierdo et al. 2005; Corsini et al. 2007; Bonnal et al. 2008).
In this article, we have characterized the molecular defect behind a mutation in the Fas/CD95 gene identified in a patient with ALPS, which leads to constitutive exon 6 skipping. The results reveal strict sequence requirements at the 3′ splice site and the existence of a rate-limiting step for exon inclusion in the recruitment of U2 snRNP after U2AF binding.
RESULTS
Strict sequence requirements in the 3′ splice site region of Fas intron 5
It has been previously shown that a C-to-G mutation at position –3 of the 3′ splice site of Fas intron 5 correlates with increased exon 6 skipping in activated T and B cells derived from a patient suffering from ALPS (Roesler et al. 2005). Because this mutation generates a tandem duplication of the 3′ splice site AG dinucleotide, the mutant will be described hereafter as AG. To confirm that the splicing defect can be exclusively attributed to this mutation and also to evaluate the possibility of a cell type–specific contribution, HeLa cells were transfected with wild-type and mutant minigenes containing Fas genomic region from exon 5–7 under a CMV promoter. After RNA isolation, the pattern of Fas splicing was analyzed by semi-quantitative RT–polymerase chain reaction (PCR) using vector-specific primers. As observed in the ALPS patient's cells, the mutation led to a dramatic switch from preferential exon 6 inclusion to quantitative skipping (Fig. 1A).
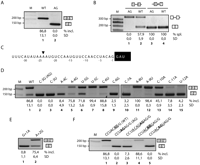
Strict requirement of RCAG/G in Fas intron 5 3′ splice site. (A) Fas minigenes encompassing exon 5 to exon 7 carrying the wild-type sequence or C-to-G mutation at position −3 of intron 5 (referred to as AG because it generates a duplicated AG dinucleotide at the 3′ splice site) found in an ALPS patient were transfected in HeLa cells. RNA was extracted, and an alternative splicing pattern was monitored by RT followed by semi-quantitative RT-PCR. The alternatively spliced products were resolved by agarose gel electrophoresis. The positions of size markers and predicted alternatively spliced products are indicated. Average percentage of inclusion (% incl.) for a minimum of three independent experiments and SD values are indicated. (B) Splicing pattern of wild-type and AG minigenes containing single introns. Analyses were as in A using primers appropriate to detect spliced and intron retention products. The vertical line in intron 6 represents a deletion that facilitates the detection of unspliced RNA but that does not affect Fas splicing or regulation (Izquierdo et al. 2005). (C) Sequence of Fas intron 5 3′ splice site region. Numbers indicate the distance from exon 6, which is represented by a black box. (D) Mutational analysis of intron 5 3′ splice site region. The indicated minigene mutants were transfected in HeLa cells and the alternative splicing pattern monitored as in A. Lanes 1 and 2 correspond to the wild-type and AG minigenes, respectively. (E) Mutational analysis of exon 6 positions +1 and +2. (F) Reconstructing an ACAGG sequence (underlined) suppresses the effects of the AG mutant and activates the mutant AG. The indicated 3′ splice site region mutants were analyzed as in D. (/) Original intron/exon boundary; (AG, bold) AG generated by the ALPS mutation; (gray boxes) acceptor site used when exon 6 is included.
To test the relative effect of the AG mutation on splicing of each of the introns flanking exon 6, minigenes containing the genomic regions between exons 5 and 6 (including the 5′ splice site of intron 6, to allow for exon definition effects), or between exons 6 and 7 (including also the 3′ 68 nucleotides (nt) of intron 5 and a deletion of intron 6 (50–1102) known not to affect Fas splicing or regulation (Izquierdo et al. 2005)) were transfected in HeLa cells, and RNA was analyzed as above. The mutation led to detection of only unspliced RNA from intron 5–containing transcripts, while the efficient splicing of intron 6 was unaffected by the mutation (Fig. 1B). Assays designed to detect intron 6–containing transcripts failed to show a higher accumulation of the unspliced transcripts in the AG mutant transcripts (Supplemental Fig. 1A). We conclude that the AG mutation leads to exon 6 skipping primarily by inhibiting splicing of intron 5, without major effects (e.g., through exon definition) on splicing of intron 6 (see also below).
To test whether other mutations can also influence exon 6 inclusion, single point mutations were introduced in this 3′ splice site region (Fig. 1C). Mutation of the C at position −3 to U (C-3U) also led to exon skipping (Fig. 1D, lane 3), indicating that the effect of the ALPS mutation is not necessarily due to the generation of a tandem of potential 3′ splice sites AG dinucleotides. Mutation of the A at position −4 to C or U also caused exon skipping (Fig. 1D, lane 4; data not shown), while transition to G had only a minor effect (Fig. 1D, lane 5), suggesting that a purine at this position is necessary for exon inclusion. Mutation of position −5 (U) to G had only minor effects, and in fact, mutation to C increased exon 6 inclusion (Fig. 1D, lanes 6,7). Mutation of position −6 (C) to another pyrimidine maintained the pattern of preferential exon 6 inclusion, while mutation of this or the C at position 7 to purine residues led to exon 6 skipping (Fig. 1D, lanes 8–10). These results suggest that the pyrimidine content of this region is important for exon 6 inclusion. Consistent with this, mutation of A to U at position −9 increased exon 6 inclusion, while mutation of pyrimidines to purines at positions −10 to −12 significantly increased exon 6 skipping (Fig. 1D, lanes 11–15). Finally, mutation of the G at exon 6 position +1 also led to quantitative exon skipping, while mutation of position +2 had only mild effects (Fig. 1E). We conclude that Fas exon 6 inclusion depends on the consensus RCAG/G at the 3′ splice site (R designates a purine; / indicates the intron/exon boundary) and that the balance between exon inclusion and skipping is finely tuned by single nucleotide differences in the pyrimidine content of the upstream sequence.
Consistent with this notion, regeneration of precisely the RCAG/G consensus around the original AG mutation led to complete reversion of the effect of the mutant and to preferential use of the newly generated AG as the 3′ splice site (Fig. 1F, lanes 1–5; data not shown).
The AG mutant inhibits intron 5 splicing in vitro and prevents prespliceosomal A complex formation
To investigate the molecular mechanism by which the AG mutant causes inhibition of intron 5 splicing, in vitro splicing assays were carried out using HeLa nuclear extracts and in vitro transcribed RNAs corresponding to wild-type and AG mutant Fas pre-mRNA sequences from exon 5 to nucleotide +25 of intron 6. Although the difference was not as dramatic as in minigene-transfected cells, the splicing efficiency of the AG mutant was significantly lower than that of the wild type. This was the case for the first catalytic step at early time points (threefold difference in the rate of accumulation of the lariat intermediate between wild type and AG), as well as for the second catalytic step at later time points (3.4-fold difference in the rate of accumulation of lariat product between wild type and AG) (Fig. 2A,B). In vitro splicing of intron 6–containing transcripts was not affected by the AG mutation (Supplemental Fig. 1B), confirming that the AG mutation affects specifically intron 5 removal.
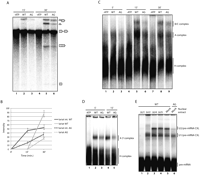
The AG mutant inhibits U2 snRNP recruitment. (A) Inhibition of pre-mRNA splicing in vitro by the AG mutant. Uniformly α32P-UTP labeled RNAs corresponding to the wild type (WT) or AG mutant Fas genomic region between exon 5 and position +25 of intron 6 were incubated with HeLa cell nuclear extracts under splicing conditions (see Materials and Methods) in the absence or presence of ATP for 15 and 30 min. RNA species were subsequently purified and analyzed by electrophoresis in 13% denaturing polyacrylamide gels. The positions of pre-mRNA, splicing intermediates, and products are indicated. (B) Quantification of lariat intermediates and products for WT and AG mutant transcripts from three independent experiments as in A. Average relative values, determined by PhosphorImager analyses, and SDs are shown. (C) Spliceosome assembly assays corresponding to in vitro splicing mixes as in A incubated for the indicated times and treated with 5 mg/mL of heparin for 10 min at room temperature prior to complex analysis by native agarose-polyacrylamide gel electrophoresis. The positions of heterogeneous nuclear RNP (H), A, B, and C complexes are shown. (D) Spliceosome assembly analysis of WT or AG RNAs corresponding to exon 6, the 3′ 68 nt of intron 5 and the 5′ 25 nt of intron 6. Analyses were as in B except that electrophoresis was carried out in 1.5% low-melting point agarose gels. The identities of the complexes are indicated. (E) Psoralen-mediated crosslinking to detect base pairing interactions between U1 and U2 snRNAs with WT or AG exon 6 splice sites. Splicing mixes as in C in the presence of psoralen (22.2 mg/mL), using mock-treated HeLa cell nuclear extracts or extracts in which U1, U2, U4, or U5 snRNAs were inactivated by RNase H–mediated digestion. The reactions were incubated for 20 min at 30°C and irradiated with long-wave UV light. RNA was isolated and analyzed by electrophoresis in denaturing 6% polyacrylamide gels. The positions of the free RNA and the U1/pre-mRNA and U2/pre-RNA crosslinked species are indicated.
Next we analyzed the step of the spliceosome assembly pathway at which the AG mutant RNA was inhibited. Lower levels of the ATP-dependent prespliceosome A and spliceosome B/C complexes were assembled on the AG mutant compared with the wild-type RNA, starting at early incubation time points (Fig. 2C). Similar results were obtained using RNAs spanning the 3′ 68 nt of intron 5, exon 6, and the 5′ 25 nt of intron 6, which assemble an ATP-dependent A3′ complex (Fig. 2D), indicating that the effect of the AG mutant does not require the presence of an upstream 5′ splice site. The same RNAs were used to monitor the interaction of U1 and U2 snRNAs with the 5′ and 3′ splice site sequences flanking exon 6 using psoralen-mediated UV crosslinking (Wassarman and Steitz 1992). As previously described (Izquierdo et al. 2005), the strong exon definition effects involved in exon 6 recognition render U2 snRNP recruitment dependent upon recognition of the downstream 5′ splice site by U1 snRNP (Fig. 2E, lane 1). The results of Figure 2E show that the AG mutation did not cause significant changes in U1 crosslinking, while crosslinking of U2 was substantially decreased (lane 6). Consistent with this conclusion, assembly of commitment complex E (containing U1 snRNP and U2AF, but not stably-bound U2 snRNP) was not affected by the AG mutation (Supplemental Fig. 2A). We conclude that the ALPS-causing mutation reduces the interaction of U2 snRNP with Fas intron 5 3′ splice region.
Initial U2AF recruitment is not affected in the AG mutant
Next we tested whether the decrease in U2 snRNP recruitment in the AG mutant could be due to decreased binding of U2AF using short wavelength ultraviolet light-mediated crosslinking and immunoprecipitation with specific antibodies (Gama-Carvalho et al. 1997). Interestingly, at early times of incubation (5 min), when clear differences in A complex formation between the wild-type and mutant AG are observed (Fig. 2C,E), no significant differences in U2AF65 crosslinking could be detected (Fig. 3A, lanes 1,2). Interestingly, however, U2AF65 crosslinking to the wild-type RNA was higher than to the mutant AG at later incubation times (lanes 3–6). These observations suggest that the AG mutation reduces the stability (e.g., by accelerating the dissociation) of U2AF65 interaction with the pre-mRNA, which may also contribute to the inhibitory effects of the AG mutation. The differential effect at late incubation times depends on ATP and is not observed at 0°C, suggesting an active process of U2AF displacement (Supplemental Fig. 2B).
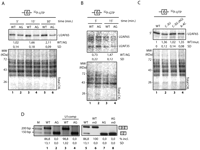
The AG mutation does not inhibit initial recognition of the 3′splice site by the U2AF heterodimer. (A) Crosslinking-immunoprecipitation of U2AF65 to assess binding to wild-type (WT) and AG mutant RNAs. 32P-uridine–labeled WT and AG RNAs comprising the 3′ 68 nucleotides of intron 5, exon 6, and 5′ 25 nucleotides of intron 6 were incubated with nuclear extracts under splicing conditions for the indicated times. After ultraviolet irradiation and RNase treatment, U2AF65 was immunoprecipitated using the MC3 monoclonal antibody (Gama-Carvalho et al. 1997). Immunoprecipitated materials were fractionated by sodium-dodecyl sulfate (SDS)–polyacrylamide gel electrophoresis. The position of RNA-crosslinked U2AF65 is indicated. The ratios of U2AF65 crosslinking to WT versus AG probes (WT/AG) at 5, 15, and 30 min and the SD corresponding to four independent experiments are indicated below the corresponding lanes. An aliquot of the products of crosslinking before immunoprecipitation was analyzed by electrophoresis and serves as loading control. (B) U2AF35 crosslinking-immunoprecipitation to WT and mutant AG. Assays were carried out as in A for the indicated times points using α-32P-guanosine–labeled RNAs. U2AF35 was immunoprecipitated using a rabbit polyclonal antibody (Zuo and Maniatis 1996). The positions of the U2AF heterodimer components crosslinked to the RNAs are indicated. The ratio of U2AF35 crosslinking to WT versus AG RNAs at 5 and 15 min and the SD for three independent experiments are indicated below the corresponding lanes. (C) U2AF65 crosslinking-immunoprecipitation to various 3′ splice site mutants. Assays were carried out as in A with the indicated mutant RNAs. Ratios and SDs correspond to three independent experiments. (D) Improving exon definition does not relieve the effect of the AG mutation. The indicated mutant minigenes were transfected and RNAs analyzed as in Figure 1. U1comp indicates a mutant minigene with enhanced complementarity between U1 snRNA and the intron 6 5′ splice site (positions −2 and −3 from the exon 6–intron 6 boundary were mutated to A and C, and positions +7 and +8 in intron 6 were mutated to A and T). The m0 mutant replaces the uridine-rich PTB-binding silencer in exon 6 (UUUGUCUUCUUCUUUU) by a random sequence (AAUGCACACUCACCAG) (Izquierdo et al. 2005).
Interaction of U2AF35 with the pre-mRNA was also analyzed using 32P-guanidine-labeled transcripts to improve the sensitivity of detection of the U2AF35/3′ splice site AG interaction. Figure 3B shows that neither the levels of U2AF35 crosslinking nor the levels of crosslinked U2AF65, which was coimmunoprecipitated with U2AF35, were lower for the AG mutant than for the wild-type RNA at 5 min of incubation (wild type/AG ratio 0.73 0.22, P-value (t-test) = 0.07), while they were slightly decreased at 15 min (wild type/AG ratio 1.47 0.12, P-value (t-test) = 0.002), as observed in Figure 3A for U2AF65.
We conclude that reduced levels of U2 snRNP recruitment in the AG mutant do not correlate with reduced levels of U2AF heterodimer crosslinking to the pre-mRNA. Interestingly, reduced crosslinking of U2AF65 was observed, even at early incubation times, for other two mutants (C-3U and A-4C) (Fig. 3C) which also led to exon 6 skipping in transfection assays, suggesting that reduced U2AF recruitment contributes to the splicing defect in these mutants.
To further explore the relationship between U2AF recruitment and the effect of the AG mutation, two additional mutants were tested. Previous work showed that increasing the complementarity between U1 snRNA and the 5′ splice site of intron 6 enhances exon 6 inclusion through exon definition effects that enhance U2AF65 binding to the Py-tract (Izquierdo et al. 2005). These bridging interactions could be prevented by the function of the splicing repressor PTB acting from a silencer sequence located in the central region of exon 6 (Izquierdo et al. 2005). Improving the strength of the intron 6 5′ splice site (U1 comp) or mutating the PTB binding site in the exonic silencer (m0) increased exon 6 inclusion (Fig. 3D, cf. lanes 1,3 and lanes 5,6). Neither of these mutations, however, increased exon 6 inclusion in the context of the AG mutant (Fig. 3D, lanes 4,8). These results are once again consistent with the notion that U2AF binding is not the rate-limiting step leading to exon 6 skipping in the AG mutant.
Suppressing the effects of the AG mutation
Previous studies have shown that the relative use of neighboring 3′ acceptor AGs depends upon their relative distance to the branch point and to each other (Chua and Reed 2001; Lev-Maor et al. 2003). To determine whether these effects were relevant in the case of the AG mutant, the distance between AGs was increased up to 5 nt. Introducing 1, 2, or 3 nt did not prevent complete exon 6 skipping (Fig. 4A, lanes 1−5). Introducing 4 or 5 nt, however, led to exon inclusion levels similar to those of the wild-type RNA (lanes 6−8), resulting in the use of the 5′-most acceptor site (as determined by sequencing of the cDNA products; data not shown). These effects are likely due to the increase in the distance between AGs because they were observed with different intervening sequences (lanes 1−8) and because computational searches (http://sroogle.tau.ac.il) failed to identify regulatory sequences that could have been spuriously introduced by the mutations that could explain the observed changes. The presence of a downstream AG plays a role in inhibition, because mutation of the 3′ AG partially relieved exon 6 skipping (cf. lanes 11 and 4), while it did not promote exon inclusion in a transcript where the more distal AG did not display an inhibitory effect (cf. lanes 10 and 6). These observations indicate that the presence of a 3′ distal AG <4 nt from the proximal AG contributes to the inhibition of exon 6 inclusion in the AG mutant.
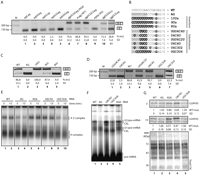
Suppression of the AG mutation by increasing the distance between AGs and by stronger Py-tracts. (A) AG-AG insertion mutants. Analysis of the indicated mutants were as in Figure 1. The first ag indicates the position of the AG created by the ALPS mutation; the second corresponds to the natural intron 5 3′ splice site. Underlined ags are those used when exon 6 is included (determined by sequencing of the spliced products). (B) The 3′ss sequence of WT, AG, and Py-tract mutants. Mutated/inserted nucleotides are in bold. (/) Intron/exon boundary. The box summarizes the conclusion from the results of parts C and D that at least six uridines are required to promote exon 6 inclusion in the AG mutant. (C) Analysis of the effects of uridine insertions in the AG mutant, analyzed as in A. (D) Analysis of mutants showing different arrangements of uridine-rich regions in the Py-tract, analyzed as in A. (E) Spliceosome assembly analysis of AG-AG insertion and Py-tract mutants, analyzed as in Figure 2D at the indicated times of incubation. (F) Psoralen-mediated crosslinking analysis for detection of U1/Fas pre-mRNA and U2/Fas pre-mRNA base-pairing interactions, performed as in Figure 2E with wild-type or the indicated mutant RNAs. (G) U2AF65 crosslinking-immunoprecipitation assays were carried out as in Figure 3A using wild-type or the indicated mutant RNAs.
Next, we evaluated the contribution of the strength and configuration of the Py-tract on the effect of the AG mutation (Fig. 4B). Inserting eight uridines and three cytidines upstream of the AGAG sequence led to a modest increase in exon 6 inclusion (Fig. 4C, lane 5). Insertion of nine or 10 uridines, however, led to quantitative exon inclusion (lanes 3,4), revealing the remarkable effect of a single additional uridine for activation of the mutant AG as a 3′ acceptor site.
Because insertion of 10 nt at the Py-tract could alter 3′ splice site recognition by increasing the distance between the branch point and the 3′ splice site, substitution mutants in the Py-tract were also generated (Fig. 4B). Substantial levels of exon 6 inclusion were observed in constructs containing six (but not four) contiguous uridines (Fig. 4D, lanes 1−5). Interestingly, however, the position of the six uridines was functionally important because a six-uridine stretch located just upstream of the 3′ splice site AG was not able to promote exon 6 inclusion, while the same length uridine stretch located further upstream induced predominant exon inclusion (Fig. 4D, lanes 5−7).
To investigate the molecular mechanisms behind the effects of these mutations, spliceosome assembly, psoralen-mediated RNA/RNA crosslinking, and U2AF65 crosslinking/immunoprecipitation assays were carried out using radioactively labeled RNAs as in Figure 3. A good correlation was found between A 3′ complex assembly, U2 snRNA psoralen-mediated crosslinking, and exon 6 inclusion (Fig. 4, cf. A, D, E, F). Specifically, wild-type Fas showed similar U2 snRNP assembly and U2 snRNA crosslinking that mutants AG6 (AGs separated out by 4 nt) and U6C4U (six-uridine stretch at a distal position from the upstream AG) (Fig. 4E, lanes 1,2,5−8, and F, lanes 1,3,5), consistent with preferential inclusion of exon 6 observed upon transient transfection of constructs harboring these mutations (Fig. 4A,D). Conversely, reduced levels of U2 snRNP assembly and U2 snRNA crosslinking were detected for the AG and the UGC3U6 (six-uridine stretch proximal to the upstream AG) mutants (Fig. 4E, lanes 3,4,9,10, and F, lanes 2,4), consistent with preferential exon 6 skipping observed in transient transfection assays (Fig. 4D).
Finally, we monitored the interaction of U2AF65 with the Py-tract at 5 and 15 min by ultraviolet-mediated crosslinking followed by U2AF65 immunoprecipitation. At 5 min, U2AF65 crosslinked efficiently with the wild-type, AG, AG6, and U6C4U RNAs. The interaction decreased in the AG mutant at 15 min, while it was maintained or even increased at this time point with the wild type, AG6, and U6C4U (Fig. 4G, lanes 1−4). In contrast, the UGC3U6 mutant, which displayed exon 6 skipping and reduced complex A assembly in vitro, showed reduced U2AF65 crosslinking at either time point (Fig. 4G, lane 5).
We conclude that the splicing defect generated by the AG (C-3G) mutation can be relieved by increasing the distance between the newly formed AG dinucleotide and the natural 3′ splice site AG, by mutating the natural 3′ splice site, or by increasing the strength of the Py-tract. The latter can be achieved with a Py-tract of six or more consecutive uridines located not immediately upstream of the 3′ splice site. The results also show that while decreased U2AF65 interaction can be correlated with decreased binding of U2 snRNP to the 3′ splice site region of intron 5 (e.g., mutant UGC3U6), decreased exon 6 inclusion in the ALPS mutation cannot be explained by decreased interaction with U2AF65, but rather by a defect in U2 snRNP recruitment subsequent to U2AF65 binding.
DISCUSSION
The consensus 3′ splice site in human introns is NYAG/R (N indicates any nucleotide; Y, pyrimidine; R, purine; /, intron-exon boundary) (Zhang 1998). Our results show that the function of the 3′ splice site of Fas intron 5, and consequently inclusion of the alternatively spliced exon 6, requires the sequence RCAG/G (Fig. 1), which differs from the general consensus (Zhang 1998) in several important features. First, a purine is required at position −4, while no clear nucleotide preference can be found at this position for the bulk of 3′ splice sites. Second, a cytidine is strictly required at position −3, while uridine can be found at this position in up to 37% of human 3′ splice sites. Third, guanidine cannot be replaced by adenine at position +1, while adenine can be found in up to 26% of human sites. The consensus CAG/G emerged as the high affinity site selected by the U2AF 35-kDa subunit in iterative binding selection experiments using the U2AF65-U2AF35 heterodimer (Wu et al. 1999). It is therefore conceivable that the strict sequence requirements at positions −3 and +1 reflect the need for tight U2AF35 binding in Fas intron 5 3′ splice site activation. While some of the mutants breaking the RCAG/G consensus decreased U2AF65 crosslinking, no differences in U2AF35 crosslinking were observed between wild-type and AG mutant RNAs in nuclear extracts (Fig. 3B). This could reflect a limitation of the crosslinking assay but also that other factor(s) specifically involved in recognition of this 3′ splice site display exact sequence requirements for positions −3, +1 (and perhaps also for the uncharacteristic need of a purine at position −4). Biochemical experiments using site-specifically radioactively labeled RNAs as well as RNA affinity chromatography, however, failed to reveal factors that differentially interact with the wild-type and AG mutant 3′ splice sites (data not shown).
While U2AF65 crosslinking was similar to the wild type and AG mutant at early time points (when differences in spliceosome assembly were already obvious), crosslinking signals were lower for the AG mutant at later times (Fig. 3A). This may reflect higher off rates of U2AF binding in the mutant or—intriguingly—the existence of a proofreading activity that displaces more readily U2AF from the mutant RNA after the initial steps of spliceosome assembly (Supplemental Fig. 2B). In this regard, the protein DEK proofreads interactions of U2AF with the Py-tract RNAs followed by a bona fide 3′ splice site AG (Soares et al. 2006) and could play a role in this process.
Alternatively, other aspects of U2AF35 or U2AF65 activities leading to U2 snRNP recruitment may display strict 3′ splice site sequence requirements in the context of Fas intron 5. Two nonmutually exclusive mechanisms have been proposed for how U2AF promotes the recruitment of U2 snRNP. Upon recognition of the Py-tract by U2AF65, its RS domain is positioned in the vicinity of the branch point region and promotes base pairing between this region and the branch point recognition site in U2 snRNA (Valcárcel et al. 1996). An interaction between the C-terminal U2AF Homology Domain (UHM) of U2AF65 and the U2 snRNP protein SF3b 155 can also contribute to U2 snRNP recruitment (Gozani et al. 1998). It is therefore conceivable that even though similar U2AF binding occurs on the wild type and on the AG mutant 3′ splice site region, the architecture of the complex formed (including conformational changes in the U2AF subunits induced upon RNA binding) differs in the mutant such that proper positioning of the RS domain near the branch point region, or proper interaction of the UHM domain with SF3b 155, cannot occur. The observation that rescue of the effect of the AG mutant by increased uridine content in the Py-tract requires at least six uridines positioned not immediately upstream of the 3′ splice site is consistent with the requirement of a special RNP structure for activation of intron 5 3′ splice site. Consistent with the notion of 3′ splice site architectures highly sensitive to mutation, in vivo evidence has been reported in Caenorhabditis elegans that single nucleotide variations can have profound effects in 3′ splice site selection (Ma and Horvitz 2009).
Although similar effects are induced by the AG mutation and by other mutants in the RCAG/G consensus, the generation of a new potential 3′ splice site is relevant for the splicing defect observed in the ALPS-causing mutation. The newly generated AG dinucleotide can be engaged in splicing reactions when the distance with the natural AG is increased by ≥4 nt or when the natural site is mutated (Fig. 4), indicating that the natural 3′ splice site prevents the use of the newly created 3′ splice site. This is another peculiarity of intron 5 3′ splice site region, because tandem AG dinucleotide arrangements frequently do not result in exon skipping but rather in activation of the mutant AG for splicing (as is the case of the hemophilia-causing mutation transmitted in certain European royal families; Rogaev et al. 2009). In addition, NAGNAG acceptors are very common in genes from a variety of organisms from land plants to humans (Hiller et al. 2004; Schindler et al. 2008). Although the use of the alternative NAGNAG sites (known as 3′ splice junction wobbling) has been argued to be stochastic (Chern et al. 2006; Sinha et al. 2009), evidence exists for functional effects of the single amino acid differences generated by the use of the alternative acceptors (Tsai et al. 2008) and for arrangements of regulatory sequences at these regions (Akerman and Mandel-Gutfreund 2006; Tsai et al. 2007).
The splicing factor hSlu7 is thought to contribute to the reorganization of the spliceosome between the first and second catalytic steps of the splicing reaction and to play a role in holding the free 3′ OH of the released upstream exon near the 3′ splice site AG to allow exon ligation (Chua and Reed 1999a). In the absence of the fidelity function of hSlu7, other neighboring AGs become activated and used for splicing (Chua and Reed 1999b). Knock-down of hSlu7 failed however to affect wobble splicing choices (Tsai et al. 2010) and also failed to relieve the inhibitory effect of the AG mutant (data not shown).
Another factor that can play a role in the selection between alternative AGs is SPF45 (Lallena et al. 2002). SPF45 regulates alternative splicing of Fas exon 6, an activity that requires interactions between SPF45 and U2AF65, SF1/BBP, and SF3b 155 (Corsini et al. 2007). Knock-down of SPF45 (as well as knock-down of a variety of other RNA binding proteins) failed, however, to promote exon inclusion in the mutant AG (data not shown).
A variety of other factors regulate Fas alternative splicing, including TIA-1, PTB, and RBM5. TIA-1 promotes exon inclusion by facilitating U1 snRNP recruitment to the 5′ splice site of intron 6 and exon definition, while PTB causes exon skipping by binding to an exonic silencer motif and interfering with exon definition (Förch et al. 2000; Izquierdo et al. 2005). The target of these regulators through the exon definition process is U2AF binding to intron 5 3′ splice site, therefore making them unlikely contributors to the splicing defect caused by the AG mutant, which is also consistent with the results of the mutational analysis of Figure 3D. RBM5 regulates exon 6 skipping by blocking splice site pairing after U2 snRNP binding, at the time of U4/5/6 assembly (Bonnal et al. 2008), an activity that therefore operates after the step blocked in the AG mutant.
Our mutational analyses show that regulation of U2 snRNP recruitment after U2AF binding can very significantly alter the balance between exon inclusion and skipping, opening the possibility that yet to be identified regulators of Fas exon 6 splicing target this important step. More generally, our results reveal that the particular configuration of sequences at this 3′ splice site is delicately tuned, being very sensitive to nucleotide replacement and positional effects. This is in contrast with the apparent flexible sequence requirements suggested by loose consensus sequences derived from comparison of a large number of 3′ splice site regions. For example, a genome-wide study in fission yeast revealed a wide array of sequence arrangements at 3′ splice sites, ranging from conventional position of the Py-tract to functional Py-tracts located 5′ of the branch point to 3′ splice site regions lacking any detectable Py-tract (Sridharan and Singh 2007). Consistent with the standard function of U2AF, the same study reported variable dependence of U2AF large subunit for intron removal.
Collectively, these considerations suggest, on one hand, that the specific RNP architecture of particular 3′ splice site regions can be finely adjusted and therefore be very sensitive to mutation, an observation relevant for splicing regulation as well as for ALPS and a variety of other genetic diseases caused by splicing defects (Buratti et al. 2006). On the other hand, they suggest that a variety of sequences, factors, and regulatory steps can contribute to 3′ splice site recognition through mechanisms that for the most part remain to be understood.
MATERIALS AND METHODS
Plasmid mutagenesis
Mutants were generated by PCR-based site-directed mutagenesis of the expression vector containing human Fas genomic sequences as described (Forch et al. 2000) using primers carrying the desired mutation, 2.5 U of TaqPlus Precision Polymerase (Stratagene) and 26 amplification cycles (for 1 min at 94°C, for 1 min at 56°C, and 7 min at 72°C). PCR products were purified (PCR purification kit, Qiagen), digested with 20 U of DpnI (New England BioLabs), and treated with 5 U of T4 DNA polymerase (Fermentas). After precipitation, ligation was carried out with 400 U of T4 DNA ligase (New England BioLabs) and transformed into heat shock– or electro-competent XL1-Blue cells. All the mutants were confirmed by sequencing.
DNA transfections, RNA purification, and analysis
Exponentially growing HeLa cells grown in Dulbecco's modified eagle's medium (DMEM), 10% FCS (fetal calf serum), and 1% penicillin/streptomycin were transfected at 50% confluence (5 × 106 cells) with 0.2 μg of plasmid DNA using ExGen 500 (Fermentas) or Fugene (Roche) following the manufacturer's instructions for adherent cell lines. Cells were incubated 24 h before RNA purification, and total RNA was isolated using RNeasy kit (Qiagen). RT-PCR was carried out as described (Forch et al. 2000). Briefly, RT was performed using 1 μg of DNase-treated total RNA and the PT2 minigene-specific primer (5′-aagcttgcatcgaatcagtag-3′) for 1 h at 42°C. Semi-quantitative PCR was then performed using GoTaq DNA polymerase (Promega) and 24 amplification cycles (for 1 min at 93°C, for 45 sec at 60°C, and for 1 min at 72°C) using oligonucleotides PT1 (5′-gtcgacgacacttgctcaac-3′) and PT2. PCR products were analyzed by electrophoresis in 2% agarose gels.
In vitro transcription of splicing substrates
Transcription templates were generated by PCR from the different plasmids, with a T7 promoter included in the upstream primer. Radiolabeled pre-mRNAs were transcribed by using ∼1 μg of template DNA in 25 μL transcription reactions containing final concentrations of 40 mM Tris-HCl (pH 7.9); 10 mM NaCl; 6 mM MgCl2; 2 mM spermidine; 0.8 mM dithiothreitol; 0.5 mM ATP, CTP, and GTP; 0.05 mM UTP; 30 μCi [α-32P]UTP; and 60 U of T7 or 42 U of SP6 RNA polymerases (Promega), for 2 h at 37°C. The transcripts were isolated by gel purification followed by phenol/choloroform/isoamylalcohol (25:24:1) extraction and ethanol precipitation.
In vitro splicing and spliceosome assembly assays
Body radiolabeled RNA (10 fmol) was incubated with 33% of HeLa cells nuclear extract, 3.3% polyvynil alcohol, 1 mM ATP, 2.7 mM MgCl2, 20 mM creatine phosphate, 4 U of RNasin (Promega), up to 9 μL with buffer D, 0.1 M KCl, 1 mM DTT at 30°C for the indicated time points. For in vitro splicing assays, the reaction was stopped by treating for 20 min at 65°C with proteinase K. The RNAs were isolated by phenol/choloroform/isoamylalcohol (25:24:1), ethanol precipitated, and loaded on 13% denaturing polyacrylamide gels. Gels were analyzed by exposing directly to PhosphorImager screens. For spliceosome assembly assays, the reactions were treated with 5 μg/μL of heparin for 10 min at room temperature and resolved on 4% acrylamide:bisacrylamide (80:1)-0.5% agarose gels in 50 mM Tris base–50 mM glycine buffer. The gels were dried and exposed to a PhosphorImager screen. Alternatively, 2.2 μL of 6× DNA loading dye (20 mM Tris-HCl at pH 7.5, 0.25% bromophenol blue, 0.25% xylene cyanol, 30% glycerol) was added to the reaction after treatment with heparin (5 mg/mL) for 10 min at room temperature. Mixtures were resolved either in agarose-polyacrylamide gels or in 1.5% low-melting agarose gels (Ecogen) in 50 mM Tris base–50 mM glycine buffer. The gels were fixed in 10% methanol–10% acetic acid for 10 min at room temperature, dried, and exposed to a PhosphorImager screen.
UV crosslinking and immunoprecipitation
32P-labeled RNAs were incubated under in vitro splicing conditions in a total volume of 27 μL. After incubation for 30 min at 30°C, mixtures were irradiated with ultraviolet light (254 nm; 0.4 J). After incubation with 1 mg/mL RNase A for 30 min at 37°C, U2AF65 and U2AF35 were immunoprecipitated with the MC3 monoclonal antibody hybridoma supernatant (Gama-Carvalho et al. 1997) and a polyclonal antibody (Zuo and Maniatis 1996), respectively. After incubating for 60 min at 4°C, 30 μL of a 50% slurry of protein A/G sepharose beads was added, and the mixture was incubated under rotation for another 60 min. Beads were sedimented by centrifugation and washed four times with 800 μL of high-salt buffer (500 mM NaCl, 50 mM Tris-HCl at pH 8.0, 1% NP-40) and once with the same buffer with 100 mM NaCl. Sedimented beads were resuspended in 4× SDS loading dye, heated for 5 min at 95°C, and briefly centrifuged, and the supernatant was loaded on 10% SDS-polyacrylamide gels. Gels were dried and exposed to a PhosphorImager screen.
Psoralen-mediated RNA-RNA crosslinking
Fifteen-microliter splicing reactions containing 80 fmol of 32P-uridine–labeled RNAs were incubated at 30°C for 20 min in the presence of 22.2 μg/mL 4′ aminomethyltrioxsalen hydrochloride (Sigma). The mixture was irradiated with 365 nm wavelength ultraviolet light for 10 min. RNAs were then treated with proteinase K, ethanol precipitated, and fractionated on a denaturing 6% polyacrylamide gel. RNase H mediated inactivation of U2 snRNP was performed as described previously (Merendino et al. 1999) by incubating nuclear extracts with oligodeoxynucleotides complementary to U2 snRNA (5′-CAGATACTACACTTG-3′), U1 snRNA (5′-CTGCCAGGTAAGTAT-3′), U4 snRNA (5′-GGGTATTGGGAAAAGTTTTCAATTAGCAATA-3′), and U5 snRNA (5′-TTAAGACTCAGAGTTGTTCC-3′).
SUPPLEMENTAL MATERIAL
Supplemental material is available for this article.
ACKNOWLEDGMENTS
We thank Robin Reed (Harvard Medical School), Joachim Roesler (Technical University, Dresden), and Adrian Krainer (Cold Spring Harbor Laboratory) for support, advice, and comments on the manuscript and members of our laboratory for discussions and input on the project. This work was supported by grants from Fundación Marcelino Botín, Fundación Alicia Koplowicz, AICR, EURASNET, Ministerio de Ciencia e Innovación, and Consolider Ingenio Program. A.C. was supported by an FPI fellowship from the Ministerio de Ciencia e Innovación.
Footnotes
-
Reprint requests to: Juan Valcárcel, Centre de Regulació Genòmica, Dr. Aiguader, 88, 08003 Barcelona, Spain; e-mail: juan.valcarcel{at}crg.es; fax: 34-9322-40899.
-
Article published online ahead of print. Article and publication date are at http://www.rnajournal.org/cgi/doi/10.1261/rna.2444811.
- Received August 31, 2010.
- Accepted November 10, 2010.
- Copyright © 2011 RNA Society








