
Latent splice sites and stop codons revisited
Abstract
The accuracy of the data we reported in an RNA Letter to the Editor earlier this year on the possible relationship between stop codons and splicing is questioned by Miriami et al. (this issue). We reply here that we see no inaccuracy in our data presentation and offer a possible explanation for their interpretation.
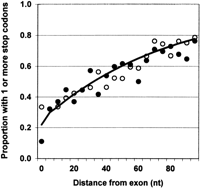
We recently reported a genomic analysis testing the model of Li et al. (2002) that in-frame stop codons (SCs) upstream of false or “latent” 5′ splice sites (LSs) were a determining factor in distinguishing false sites from true sites (Zhang et al 2003). We found no evidence for an increased frequency of stop codons upstream of such LSs, arguing against this model (Zhang et al. 2003). Miriami et al. (2003), in their response to that report, claim that we erred in plotting our data. We see no errors in our graph, and believe these authors misconstrued what was plotted. We reproduce the graph here and more fully explain what was plotted (Fig. 1).
We reasoned that if in-frame stop codons acted as general signals to ignore a downstream LS, then they should always be found between the real 5′ splice site and the LS. If this model is not correct, then the frequency of in-frame SCs should be not greater than that expected by chance. We chose to plot these data as a function of the position that the LS occupies downstream of the exon boundary, because both predictions approach the same frequency of one at distances greater than ~100 nt.
The X-axis in the figure represents introns grouped by the distance at which their most 5′ LS is found (in windows of five). The number of introns in each class ranged from 21 to 82. The Y-axis represents the proportion of each intron class that have at least one in-frame SC upstream of the LS.
The solid line represents the expected proportion of introns in each class that have at least one SC upstream of the LS if this occurrence happened by chance. To make this calculation, we divided the 100-nt region into two components. The first 6 nt of all introns comprise part of the splice site itself, which has a high probability of harboring an SC because of the URA embedded in its consensus sequence AG/GURAGU. Based on the matrix of bases underlying this consensus sequence, we calculated that the probability of an SC sequence occurring by chance is 0.74. We divided this number by 3 for the three possible reading frames to obtain this probability for an in-frame occurrence (0.247). Based on a Poisson distribution, the expected frequency of introns having an in-frame SC within the splice site is 1 − P0, where P0 = e−0.247.
The second component represents the probability of a stop codon occurring within the remaining 94 nt of this 100-nt flank. We used the Poisson distribution to calculate the fraction of introns in different distance classes expected to contain at least one in-frame SC. We calculated P0 based on the mean SC frequency per triplet (m) in the regions 100 nt downstream of all real introns, regardless of the presence of an LS. For this calculation, the 92 triplets from positions +7 to +98 were considered. Thus, here P0 = e−m(L − 6)/3, where m is 0.038 per triplet and L is the distance from the exon border to the lower limit of the LS class. These two components were combined to obtain the expected fraction (E) of introns harboring at least one SC, as E = (1 − e−0.247 × e−m(L − 6)/3).
The observed data are shown by the solid circles, which represent the actual fraction of each intron class that contains at least one SC upstream of the LS. The open circles represent, for comparison, the same data for the two out-of-frame stop codons (the average for these two frames). There is no apparent difference between in- or out-of-frame SCs, and the points for both follow the expectation based on chance. We thus stand by our original conclusion that these data do not support a general role for stop codons in introns acting as negative splicing signals.
The two left-most points, corresponding to an LS within 1–5 nt (for instance, in the splice site itself) do deviate from our expectation based on chance: in-frame SCs are less than expected and out-of-frame SCs are greater than expected. This result can be anticipated from the tendency of exons to end in translation phase 0 (about one-half rather than one-third of exons end between codons), putting the URA at positions +2 to +4 of the consensus out of frame.
The confusion may have arisen from the fact that we based our estimation of the probability of SCs occurring within the splice site by chance on the weight matrix (as stated in our paper), whereas Miriami et al. (2003) apparently assumed that we used the actual frequency of in-frame SCs, as they did in a related but different type of analysis.
The occurrence of nonsense codons in introns.
Footnotes
-
Article and publication are at http://www.rnajournal.org/cgi/doi/10.1261/rna.5211704.
- Copyright 2004 by RNA Society








